新版 日本人になった祖先たち
茂木誠さん(日本人はどこから来たのか?【CGS 茂木誠 超日本史 第1回】< https://youtu.be/fKc35G52wWg > )と竹田恒泰さん(【竹田学校】歴史・岩宿時代編④~ミトコンドリアからわかった日本人の起源~< https://youtu.be/_H3T6EHTaIE >、 【竹田学校】歴史・岩宿時代編⑤~Y染色体からわかった日本人の起源~< https://youtu.be/Qm41EiUhdfs >)のユーチューブ動画でDNA分析による人類・日本人の成り立ちを見ました。
せっかくなので研究者の解説も読んでみようと思い本書を手にしましたが、とても参考になりました。
本書はDNA分析に関する解説だけでなく、著者の憲法観も書かれていますが、その部分の記述は賛同できかねる内容でした。
また、著者は研究を通じてアイヌ協会とも親しいようですが、アイヌ民族の認定にもDNA鑑定を使うことを強く勧めて欲しいと思いました。
それらの個所は緑色の文字で示しました。
篠田謙一さんの「新版 日本人になった祖先たち」を紹介するために、以下に目次や目を留めた項目をコピペさせていただきます。
興味が湧いて、他も読んでみたいと思ったら、本書を手にしていただければと思います。

目次
はじめに 3
私たちのルーツを求めて ―― 人類学の目指すもの/DNAと人類学
第1章 遺伝子から私たちのルーツを探る 17
遺伝子はどのように受け継がれるか/DNAの変異を利用する個人鑑定/遺伝子から見た「私」/祖先をさかのぼるということ/DNAで系統をたどる/分子人類学の誕生/「スモール・イズ・ビューティフル」・ヒトのミトコンドリアDNA解析の歴史/父系から息子に受け継がれるY染色体DNA/Y染色体DNA分析の利点と難点/核DNAの解析法/遺伝子の分布からヒトの移動を考える/拡散と移動の諸相
第2章 アフリカから世界ヘ ―― DNAが描く新人の拡散 51
現生人類は4グループに分かれる/多くの突然変異を持つアフリカ人/Y染色体から探る人類の共通祖先/核ゲノムが語るアフリカ集団/人類の始まりの姿/最初にアフリカを旅立った集団/出アフリカの2つのルート/なぜアフリカなのか
第3章 DNAが描く人類拡散のシナリオ 71
拡散の跡を探る/ミトコンドリアDNAハプログループから見た人類の分岐/歴史を再現することの難しさ/核ゲノムで再現するヨーロッパの歴史/ヨーロッパにおける狩猟採集民の系統/ヨーロッパの農耕民/ヨーロッパ人の遺伝子を一変させた牧畜民の流入
第4章 アジアヘの展開 89
南アジアの状況/古代ゲノム解析が明らかにするインドーヨーロッパ集団の成立/東アジアと東南アジア ―― 南北に分かれる世界/東南アジアと東アジアの集団の特徴/中央アジア ―― シルクロード、北の回廊/新大陸へ渡った人たち ―― 南北アメリカ/アメリカ先住民はどこから来たのか/ミトコンドリアDNAから考えるアメリカ先住民の由来/古代アメリカ人のゲノム解析/ナスカの子供ミイラに宿る遠い旅路
第5章 現代日本人の持つDNA 113
日本人の持つミトコンドリアDNA/各ハプログループの起源地と拡散の推定
ハプログループD ―― 東アジアの最大集団
ハプログループB ―― 環太平洋に広がる移住の波
ハプログループM ―― 7日本の基層集団を生む系統
ハプログループA ―― 北東アジアに展開するマンモスハンターの系譜
ハプログループG ―― 北方に特化する地域集団
ハプログループF ―― 東南アジアの最大集団
ハプログループN9 ―― 南北に分かれるそのサブグループ
プログループM8a ―― 中原に分布する
ハプログループC ―― 中央アジアの平原に分布を広げる
ハプログループZ ―― アジアとヨーロッパを結ぶ人々
日本人のY染色体DNA/日本固有のハプログループ/核ゲノムに現れた現代日本人の地域差/集団の変遷について
第6章 日本人になった祖先たち 149
日本人起源論の系譜/二重構造説の問題点/古人骨のDNA分析の歴史/日本の古人骨/DNAから見た縄文人/ミトコンドリアDNAが示す縄文人の地域性/縄文人とは誰なのか/現代人に受け継がれる縄文人のDNA/縄文人のY染色体DNA/縄文人の核ゲノム解析/多様な弥生人/弥生人のDNA/縄文人と弥生人の混合のシナリオ歴史時代の日本人
第7章 南北の日本列島集団の成り立ち 195
多民族集団としての日本列島の歴史/旧石器時代の琉球列島集団/沖縄の縄文人(貝塚前期)のDNA/弥生時代からグスク時代までの沖縄/グスク時代のDNA/北海道先住民の成立史
第8章 DNAが語る私たちの歴史 213
国家の歴史を超えて/家系とDNAのアナロジー/核ゲノム分析の意味するもの/DNAのネットワークとしての私たちの社会/これからの社会と私たちのDNA
参考文献 226
あとがき 234
私たちのルーツを求めて ―― 人類学の目指すもの
北海道の最北端に位置する礼文島には10力所以上の縄文時代の遺跡があります。そのなかのひとつの船泊遺跡では、昭和初期に初めて学術的な発掘調査が行われ、その後も小規模な考古学的調査が行われてきました。1998年には町の教育委員会によって建設工事に伴う大規模な発掘調査が行われ、縄文時代後期の住居跡や作業場跡や墓などの遺構と、土器、石器、骨角器、貝製品など大量の文化遺物とともに28体の人骨が発見されています。私たちの研究チームは、その中の2体の人骨からDNAを抽出し、次世代シークエンサという、大量のDNA配列を一度に読み取ることのできるマシンを使って解析を行いました。その結果、そのうちの1体、40歳代の女性(23号人骨)ではその全ゲノムを現代人と同じレベルの精度で決定することができました。
ゲノムというのは、「その人の持つ遺伝子の全体を含むDNA配列」を指します。ですからゲノムが完全に読めると、その人物に関するさまざまな情報が手に入ることになります。用語や考え方については第1章以降で詳しく説明しますので、そちらを読んでいただいてからの方が理解は深まると思いますが、ここでは23号人骨についてわかったことの概要を述べます。
彼女が女性であることは骨の形の観察からもわかっていましたが、DNAでも証明されました。男性の持つY染色体DNAが検出されなかったのです。また血液型はA型でした。彼女が父と母から受け継いだDNAの配列同士を比較してみると、互いが非常によく似ていることがわかりました。詳しい解析によって、長い間少人数の集団内での婚姻が繰り返されていたこともわかりました。この遺跡からは南方産のイモガイで作られたペンダントや、新潟県糸魚川産の翡翠(ひすい)、さらにはシベリアで作られたものと同じタイプの貝玉のアクセサリーなどが見つかっており、礼文島の縄文人が広い地域と交流を持っていたことが示唆されています。このような遺物の交流範囲の大きさから、人々の通婚圏も広がっていたように想像されるのですが、それほどでもなかった、ということになります。実際のところ、ヒトとモノの動きを同一視することはできず、両者が一致する保証はありません。この結果は、ヒトの移動に関しては遺物ではなく、遺伝子を検討しなければならないことを示しています。
 さらに、彼女の姿形に関わる遺伝子の特徴から、人骨の形態からはわからないさまざまな顔の特徴を知ることもできました。図0‐1は、その情報をもとに復元されたこの女性の復顔像です。髪は縮れており、虹彩の色は茶色でした。肌の色は濃く、シミのリスクがあることもわかりました。お酒にも強い遺伝子を待ち、身長は高くならないという結果も得られています。実際の骨の観察から、彼女の身長は縄文人のなかでも小さいことがわかっていましたから、その結果は一致しています。成人の身長は、成長期の栄養状態にも依存しますが、この人はもともと高身長になることはできなかったようです。姿形や体質とDNAの関係に関する研究は、いまも猛烈な勢いで進んでいますから、将来的にはDNAの変異に関する新たな知見が付加されていくことで、この復顔像も姿を変えていくことになるでしょう。それは同時にゲノム研究の進歩を示すものになるはずです。
さらに、彼女の姿形に関わる遺伝子の特徴から、人骨の形態からはわからないさまざまな顔の特徴を知ることもできました。図0‐1は、その情報をもとに復元されたこの女性の復顔像です。髪は縮れており、虹彩の色は茶色でした。肌の色は濃く、シミのリスクがあることもわかりました。お酒にも強い遺伝子を待ち、身長は高くならないという結果も得られています。実際の骨の観察から、彼女の身長は縄文人のなかでも小さいことがわかっていましたから、その結果は一致しています。成人の身長は、成長期の栄養状態にも依存しますが、この人はもともと高身長になることはできなかったようです。姿形や体質とDNAの関係に関する研究は、いまも猛烈な勢いで進んでいますから、将来的にはDNAの変異に関する新たな知見が付加されていくことで、この復顔像も姿を変えていくことになるでしょう。それは同時にゲノム研究の進歩を示すものになるはずです。
従来の人類学の研究では、発掘された人骨の間の血縁関係や他の集団との系統関係などは、骨のさまざまな部分を計測したり、骨に現れた特徴などを調べて比較することによって推定してきました。日本の人類学者は、明治以来一世紀以上にわたって発掘人骨に対してこのような解析を行ってきたのです。現在では、私たち日本人は、船泊遺跡に埋葬されていたような縄文人と、その後の弥生時代になって大陸から渡ってきた渡来系弥生人と呼ばれる人たちの混血によって成立したと考えられていますが、このような学説も基本的には骨や歯の形態学的な調査研究の結果、導き出されたものなのです。
一方、骨の形態は、遺伝的な要因と環境要因が複雑に絡み合って決定されるので、系統や血縁関係を調べる場合には骨形態に現れる遺伝的な要素を注意深く読みとる必要があります。しかしながら骨形態の遺伝様式については不明の部分も多いので、骨の形態学的な調査から導き出された結論を評価することは、たいへん難しいのです。高身長になる遺伝子を持っていても、成長期の栄養が足りなければ大きくなることができないという事実からもそれは明らかです。骨の形態学的な研究から得られた結論と歯の形の研究から導かれた結論が異なるものになる場合もあります。これに対し、古人骨に残された遺伝子の本体であるDNAを直接解析することができれば、系統や血縁といった問題に対し比較にならないほど精度の高い情報を得ることができると予想されます。
私たちのDNAには、私たち白身の生物としての歴史が書き込まれています。そのことはもう半世紀ほど前にはわかっていたのですが、最初はそれを解析する手段がありませんでした。しかし、1970年代に始まる分子生物学の爆発的とも言える発展は、ついにそのことを可能にしたのです。現在では古人骨に残るDNAの分析すら、日常的に行われるようになってきました。
本書では、主として私か研究しているミトコンドリアDNA分析と、本書を刊行した後に付け加わった核のDNA分析によって明らかになってきた、私たち日本人の成り立ちについて紹介しようと思います。まず、現代人のDNA分析からわかってきたことを説明し、その後で、冒頭で紹介した船泊縄文人などの古代人のDNA分析の結果が教えてくれることを説明しましょう。
日本人の持つミトコンドリアDNA
日本人の成り立ちを考えるために、まず最初に私たち日本人はどのようなミトコンドリアDNAを持っているかを見ていきましょう。図5-1は、日本人の持つハプログループの割合を示したものです。これは、これまでに調べられた日本人データ1000以上を使って描かれたものですから、現在日本人が持つほぼすべてのハプログループを網羅しているはずです。実際には調査していない私たちひとりひとりも、このどれかのハプログループを持っていると考えてよいでしょう。このデータが、ミトコンドリアDNAから日本人の由来を考える基本的な資料になります。
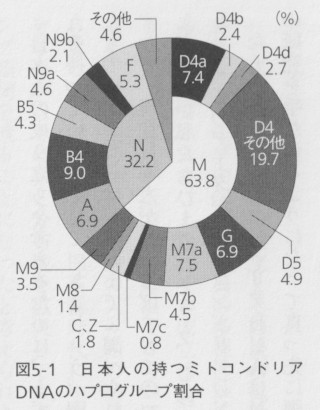
たとえば私のミトコンドリアDNAはN9aというハプログループに属しています。自分でこの研究を手がけるようになって真っ先に調べてみました。ミトコンドリアDNAは母から子へと受け継がれていきますから、私の母方の祖先はこのN9aの系統を持っており、それをさかのぼっていくと人類の共通祖先までたどり着くはずです。それが私の母系がたどってきた道なのです。
ですから日本人の由来を考えるとき、今、日本に存在するすべてのハプログループの系統を個別に調べていけば、その総体が日本人の起源、ということになります。こう書くと、それぞれのハプログループの歴史がわかっても、そもそも自分自身の持っているハプログループがわからないと、自分の由来がハッキリしないのではないか、と思う方もおられるかもしれません。しかしそれは誤解なのです。最初に説明したように母から子供に渡されるミトコンドリアDNAと、父から息子に受け継がれるY染色体の遺伝子を除く大部分のDNAは両親から受け継いでいます。たとえば私の父のミトコンドリアDNAのハプログループはAですが(これもかつて調べてみました)、これは私に伝わっていません。しかしハプログループAのたどった道も私の由来の一部のはずです。ミトコンドリアDNAのハプログループを婚姻の条件にする人はいないでしょうから、基本的に祖先における婚姻は、ハプログループに関してはランダムに行われていると考えられます。ですから実際には不可能でしょうが、仮に自分の祖先を数百人選び出して、それぞれのハプログループを調べて頻度を計算すれば、今の日本人集団が持つハプログループの割合に近いものになると思います。自分白身を構成するDNAは他の日本人と同じような経路をたどって、自分のなかに結実しているのです。そして、それぞれのハプログループのたどった道は、私たちの持つすべてのDNAのどれかがたどった道であり、その末端部に今の私たちがいるのです。こう考えると、現在の日本人が持つミトコンドリアDNAハプログループのすべてを知ることが、日本人の成立の過程を解き明かすだけでなく、私たち個人の由来について知ることになるということがおわかりでしょう。
これから、日本人に見られるハプログループのそれぞれについて、どこで生まれ、どのような経路を通って日本に入ってきたのかを、現在利用できるデータをもとに見ていくことにします。
各ハプログループの起源地と拡散の推定
それぞれのハプログループの起源地と拡散の様子をどのようにして推定するのか、最初に説明しておきましょう。簡単に言うと、あるハプログループの占める割合が他の地域よりも多くて、しかも内部の変異をたくさん持っているところを起源地と考え、そこからの拡散の様子は、人口に占める割合と変異の減少をトレースすることによって描き出すのです。それぞれのハプログループの起源地では、そのハプログループが人口に占める割合が他の地域よりも多く、かつそのハプログループのなかに含まれる変異の数が他の地域よりも大きいはずです。起源地では、他の地域に比べて、そのハプログループが誕生してから長い時間が経っていますから、それだけ内部に多くの変異が蓄積されているはずだからです。最初にアフリカ人のなかにミトコンドリアDNA全体の変異がたくさん蓄積していたことを根拠に、アフリカが人類発祥の地であると推定しました。同じことをそれぞれのハプログループの内部で想定するのです。これは他の動物や植物の起源地を推定する場合にも用いられている方法です。こうして作られたのが前掲の図4-4なのです。
もちろんハプログループの誕生は数万年前のできごとですから、誕生時点とそれから後のできごとを現在の分布だけから推定することには、そもそも無埋があります。特に歴史時代の人類集団の移動は複雑で、起源地から集団全体がいなくなってしまった場合もあったでしょう。ですから、この方法で描かれた人類拡散のシナリオは、本来、考古学や化石人骨の研究など、他のさまざまな証拠によって検証していかなければなりません。特に古人骨のゲノム解析は必須の作業になりますが、ここではとりあえず現時点でミトコンドリアDNA分析が描き出す、日本人を構成する各ハプログループの誕生と拡散の様子について見ていくことにします。
ハプログループB ―― 環太平洋に広がる移住の波
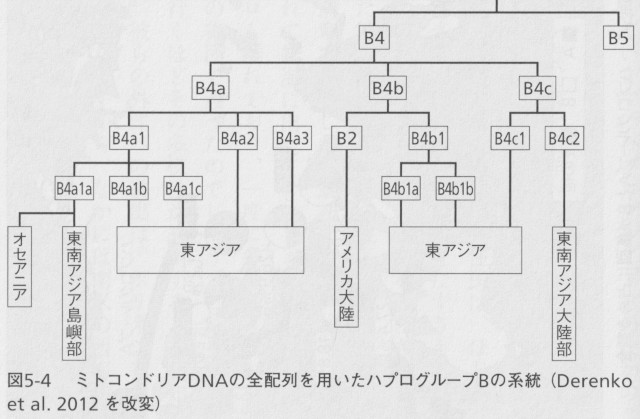
日本人のおよそ7人にひとりが該当する第2のグループがハプログループBです。前章でも述べましたが、このハプログループもアメリカ先住民で最初に見つかったものです。他のグループのように特定のDNA配列が変化しているのではなく、ミトコンドリアDNAの特定部位のDNA配列9つ分が欠損しているのが特徴です。ミトコンドリアDNAの正確な数は1万6568塩基対ですが、ハプログループBに属する人たちは1万6559塩基対しかないことになります。この9塩基対部分を含んだDNA断片をPCR法という方法で増幅してやると、比較的簡単にその有無を調べることができます。したがって、DNAの配列を読み取るのが難しかった時代には、この部分を対象にした多くの研究がなされました。その後、さまざまな集団の研究によって、この9塩基対の欠損はハプログループBの系統だけではなく、世界のいろいろな地域で独立に何度も起こったことがわかってきました。しかし現在では東アジアや新大陸で見られる9塩基対欠損は、ほとんどすべてがハプログループBの持つ変異であることも確認されています。現在、そのサブグループとしてはB4とB5を区別していますが(図5-4)、ここでは私たち日本人にもっとも多いB4について考えることにします。
ハプログループBは、およそ4万年前に中国の南部で誕生したと推定されています。このハプログループはハプログループRから分岐しますので、おそらくインドから東南アジアに拡散したハプログループRの集団のひとつから生まれたのでしょう。現在、東アジアでゲノムが決められているもっとも古い人骨は、北京の近郊にある田園洞から発見された4万年ほど前のものです。この人物の持っているハプログループがBでした。この時期にすでにかなり広い地域に広がっていたのかもしれません。図5-5はこのグループの各集団に占める割合を示したものです。誕生の地である中国南部から東南アジアにかけて人口に占める割合が大きくなっていますが、それ以外にも南米の山岳地域や南太平洋の集団に多いことがわかります。ただし、ハプログループBはこの2つの地域に同時期に拡散したのではなく、異なる時期にまったく別のルートを通って進出しました。
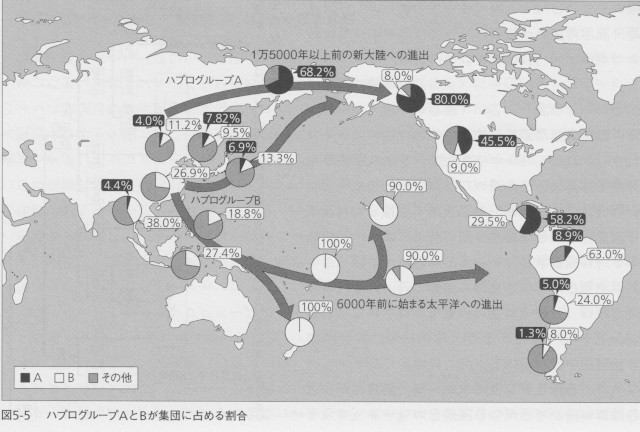
アメリカ大陸に見られるB2は、3万3000年前というB4のグループのなかでも比較的誕生時期が古い、B4bというサブグループのなかから派生しています。ハプログループBのなかのこの集団は、大陸の沿岸地帯を伝ってベーリンジアに到達し、しばらく停滞した後に新大陸に入って、そこから海岸伝いに南下したと考えられます。
一方、南太平洋への展開は約6000年ほど前のできごとですので、歴史的に見ればごく最近のことになります。こちらは、1万年ほど前に誕生したB4aのサブグループが主体となりました。現在の南太平洋の先住民は、ほとんどすべてがこのハプログループを持っています。中国南部もしくは台湾から農耕をたずさえて東南アジアの海岸地帯へ展開した集団の主体をなしたのが、このハプログループに属する人たちだったと考えられています。メラネシアの海岸地域に到達した彼らは、やがて遠洋航海の技術を身につけて、南太平洋の島々に進出します。その際、彼らは前章で説明した、最初に東南アジアに到達したハプログループPやQに属する人たちの一部も連れて航海に出たことが、その分布からわかります(前章の図4‐3)。これらの島々は互いが数百キロも離れており、一度に多量の人間が移住することは不可能です。外洋性のカヌーを使って少数の集団で渡ったのでしょう。これらの島々でミトコンドリアDNAの多様性を見ていくと、先に行くほどその多様性が減少することが知られています。
彼らの外洋への進出は、ポリネシアの島々を征服して終わりますが、そのすぐ先には南米大陸がありますから、なかには南米の西海岸にたどり着いた人たちもいたと思います。アンデス原産の植物のなかには、ヨーロッパ人が南太平洋に到達する以前にポリネシアの人々の間に伝わっていたものもあります。古代におけるアンデス住民とポリネシア人の遭遇があったことは間違いありません。そのアンデスの人たちもやはりハプログループBを主体とする人たちでした。もちろんお互いに気がつくことはなかったでしょうが、彼らは4万年前の中国南部に共通の祖先を持ち、一方は北へ、他方は南へ拡散した集団の子孫でした。数万年の時をへだてて再会を果たした親戚同士だったのです。最初の遭遇がどのようなものだったか想像することもできませんが、ミトコンドリアDNAの拡散の歴史から見れば、非常に劇的なものだったのです。
日本人のY染色体DNA
以上見たように、日本人の持つミトコンドリアDNAには数多くのハプログループが存在していたのに対し、Y染色体ではC、D、Oと呼ばれる3つの系統が人口の90%以上を占めています(図5-9)。これ以外のハプログループとしてはQとNの系統が認められているだけで、比較的単純な構成を
しているようにも見えます。ただしそれぞれのハプログループはさらに細分化されていますので、詳しく見
ていけばミトコンドリアDNAと同様、複雑な構成をしていることがわかります。それぞれのグループにつ
いて見ていくことにしましょう。

ハプログループOは、日本人の男性人口の約半数を占める最大のグループです。このグループは20以上のサブグループに細分化されていますが、日本列島に分布するのはO1b2とO2と呼ばれる系統です。日本人を対象にしたいくつかの調査がありますが、いずれも人口比でO1b2は30~40%、O2は20~30%程度を占めると報告されています。ハプログループOの分布は、東アジアやオセアニアの全域に広がっていますが、日本に分布するサブグループの分布に限ってみると、O1b2が朝鮮半島や華北地域に分布しているのに対し、O2は華北から華南にかけて広がっているようです。
ハプログループCには10種類程度のサブグループが定義されていますが、そのなかで日本に存在するのはC1aとC2と呼ばれる系統です。ハプログループCが人口に占める割合は、本土日本でおおむね10%程度ですが、北海道のアイヌの人たちでやや多いという報告もあります。もっとも、このアイヌの人たちのデータは全部で16人分のサンプルから得られた結果ですので確定的な話ではありません。もう少し例数を増やさないと比較データとしては使えないでしょう。
ハプログループCは、東アジア、オセアニア、オーストラリア、シベリア、それに南北アメリカ大陸に広く分有しています。このうちサブハプログループであるC2は、沿海州の先住民集団やモンゴルの集団に非常に高い頻度で認められ、南に行くにしたがってその頻度を減少させていきます。これに対してC1はインドネシアを中心とする地域に高頻度で認められます。この特徴的な分布から、C2の系統は北から、C1aの系統は南から日本に入ってきたと推定されます。
ハプログループDは、Y染色体のDNAにYAP+と呼ばれる余分なDNA配列が挿入されたタイプです。ハプログループEも同様の変異を持っています。ミトコンドリアDNAには9塩基対が欠損するハプログループBがありましたが、Y染色体のハプログループDやEはそれとは逆に余分なDNAが付け加わったものということになります。ハプログループEはアフリカやユーラシア大陸の西部に分布し、私たちとは直接関係がないグループですが、ハプログループDの方は日本で多数の人口を占めており、私たちにとって重要なハプログループです。このハプログループDには今のところ6つのサブグループが報告されていて、私たちが持つのは、そのうちのD1bと呼ばれるサブグループです。これまでの報告では地域によって多少のばらつきかおりますが、だいたい日本人男性の30~40%がこのハプログループを持っています。

これまで調査が行われている日本と近隣集団のY染色体とミトコンドリアDNAのハプログループ頻度をグラフにしてみました(図5-10)。このグラフを見るとすぐに気づくと思いますが、日本と朝鮮半島、中国(台湾の漢民族)で大きく違っています。その原因は、ハプログループDの頻度にあることは明瞭です。日本の近隣集団では、ハプログループDをこれだけの高頻度で持っている集団はありません。このハプログループの起源を考察するために、もう少し検索の範囲を広げて考えてみることにしましょう。
ユーラシアの東部、北東アジアの各集団には低頻度ながらD1bを見いだすことができます。またこのハプログループの近縁のハプログループD1aはチベットで人口の50%近くを占めていることも知られています。この分布は、もともと北東アジアに広く分布していたこのハプログループが、その後中国を中心とした地域で勢力を伸ばしたハプログループOの系統によって周辺に押しやられてしまった結果のように思えます。日本やチベットは海や高い山によってへだてられたので、このハプログループが高頻度で残ったのかもしれません。
この3大グループ以外のハプログループであるNとQは、いずれもアジアの北方の集団に分布しています。特にハプログループQはアメリカ先住民にも分布しており、シベリアで生まれて新大陸と北アジアの各地に広がったと推定されています。Nの系統は北欧やウラルの集団との関連性が知られており、その分布はミトコンドリアDNAのハプログループZを想起させます。どちらのハプログループも北方系の要素と言ってよいでしょうが、人口に占める割合は1%程度で、日本人においてはきわめてマイナーなグループです。
日本人起源論の系譜
前章では現代日本人の持つDNAの特徴について解説しました。この列島の内部に見られる遺伝的な特徴は、集団が成立する過程のなかで生じたものと考えられますから、それがどのように形成されたのかを考えることは、日本列島集団の成立の経緯を明らかにすることにつながります。本章ではこの問題に、古代人のDNA解析によって得られたデータを加えて説明していきますが、その前に、従来の人骨研究から導かれた日本人の起源論について紹介しておきましょう。
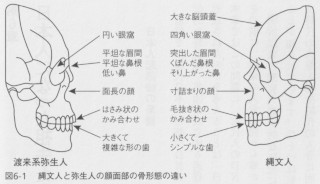
明治以来の形質人類学的な研究によって、日本列島集団の姿形には2つの大きな特徴があることが知られています。ひとつは形質には時代的な変化があるということで、具体的には、縄文時代の人骨と弥生時代の人骨に明確に認識できる違いが認められることを言います(図6-1)。ただし、この場合の縄文人というのは、今からおよそ5000年前の縄文時代中期以降の、主として関東以北の太平洋岸の貝塚に埋葬された人骨を指し、弥生人は北部九州の甕棺に埋葬された、いわゆる渡来系弥生人を指していることには注意する必要があります。実際には両者の違いは、時代的な要因の他に地理的な変異も考慮する必要があります。
2つ目の特徴は、現代の日本列島には形質の異なる集団が存在しているということで、こちらは北海道のアイヌ集団と本州を中心としたいわゆる本土日本人、そして琉球列島集団には、姿形に区別しうる特徴があるということを指しています。特に琉球集団とアイヌの人々は見た目が似ており、本土日本人との差が際立っています。このことは実感として納得できる人も多いでしょう。このような形質の違いについて、日本人の成立史の中でどのように説明されてきたかを以下に見ていきます。
今日の日本人起源論につながる議論は、日本に近代的な学問が本格的に導入された明治時代にさかのぼります。ただし明治時代の人類学は考古学や民族学を包含した学問体系で、そこで議論される日本人の起源論もこれらの分野を横断したものでした。この頃の起源論は、記紀の記述などにも影響され、基本的には人種の交替によって日本人が成立したと考えていました。先住民の住んでいた日本列島に大陸から進んだ文化を待った人たちが渡来して、国家を築いたと考えていたのです。そのようななか、大正から昭和にかけて日本各地で多数の人骨が発掘されたことによって、もっぱら発掘人骨の形態学的な研究に基づいた起源論が提唱されるようになります。その中心となったのが京都帝国大学医学部教授だった清野謙次と後に東京帝国大学の初代人類学科の教授になった長谷部言人でした。清野は現代日本人とアイヌ、石器時代人(縄文人)の人骨の比較から、日本石器時代人は日本人でもアイヌでもなくひとつの独立した集団で、これが後に周辺から渡来した人々と混血して変化したという仮説、いわゆる混血説を提唱しました。これに対し長谷部は、石器時代人と現代人の骨格形態の違いは時代変化によって説明できると考える変形説を唱えたのです。なお、この変形説の背景には、生物は進化するというダーウィンの進化論があります。長谷部の学説は日本人の形成の問題に生物学的な視点を初めて持ち込んだものだったのです。そしてこの2つの説は、その後の日本人起源論の大きな潮流となって、後継者に受け継がれることになりました。
第二次世界大戦後、混血説は九州大学の金関丈夫によって唱えられた渡来説へ受け継がれ、変形説は東京大学の鈴木尚に引き継がれることになりました。現在とは違って1980年代までは鈴木の変形説が定説と考えられていました。鈴木の研究は、それまで縄文・弥生時代の人骨だけを使って行われてきた起源論の研究に対し、その後の歴史時代の人骨をも網羅するもので、人類学者にとって大きな説得力を持つものだったのです。
日本人の骨格は歴史上2回、大きく変化します。1回目は縄文から弥生時代にかけて、2回目は江戸から明治にかけてです。この2回の画期は、いずれも日本人の生活様式が大きく変わった時期でした。前者は狩猟採集社会から農耕社会への移行、後者は西洋文明の受容です。明治時代に大量の移民はなかったのにもかかわらず日本人の体型は大きく変わったわけで、その状況を目の当たりにしていた研究者たちは、縄文・弥生移行期における変化も変形説で説明することを容易に受け入れることができたのです。またその背景として、戦後の日本に定着した日本人の単一民族神話が、この説の受容に大きく影響したことも見逃せません。第二次世界大戦後は、日本では有史以来同じ集団が存続し続けたという考え方が支配的で、渡来による混血という考え方をする人は少数だったのです。
しかし1980年頃までに、九州大学や長崎大学の解剖学教室によって北部九州地域で多数の人骨が発掘され、この地域の集団の特徴が詳しく調べられたことで状況は変化します。その結果は混血説を強く支持するもので、縄文人から弥生人への変化を変形説で説明するのには無理があることを多くの研究者が感じるようになったのです。そして現在では埴原和郎によって提唱された、旧石器時代人につながる東南アジア系の縄文人が居住していた日本列島に、東北アジア系の弥生人が流人して徐々に混血して現在に至っているという「二重構造説」が、主流の学説となっています。
この学説は、列島内だけではなく東アジアの集団の成立も含めた視野の広いもので、おおよそ以下のような集団形成のシナリオを想定しています。まず旧石器時代に東南アジアなどから北上した集団が日本列島に進入して基層集団を形成し、彼らが縄文人となります。一方、列島に入ることなく大陸を北上した集団は、やがて寒冷地適応を受けて形質を変化させ、北東アジアの新石器人となったと考えるのです。弥生時代の開始期になると、この集団の中から朝鮮半島を経由して、北部九州に稲作を日本にもたらすものが現れます。それが渡来系弥生人と呼ばれる人々です。つまり、縄文人と渡来系の弥生人はそもそも由来が異なるので、姿形に違いがあるということになります。弥生時代以降、大陸から渡来した人々は、金属器と水田稲作を全国に広めていくことになるのですが、その過程で在来の縄文人と混血していくことになったと考えています。歴史時代を通じてこの混血は進みますが、稲作が入らなかった北海道と、北部九州からおよそ2000年おくれて、10世紀頃になってようやく稲作が入った南西諸島では、縄文人の遺伝的な影響が強く残ることになります。つまり両者の見た目の類似性は、縄文人の影響であると考えるのです。この二重構造説は、列島内部に見られる時間的・空間的な形質の違いを、基層集団と渡来した集団の関係というひとつの視点で説明しているところに特徴があります。
埴原の二重構造説は、最新の統計技法やコンピュータによるシミュレーションを駆使し、大量の渡来人が存在した可能性を示唆したものでした。渡来人の数が100万人にもなりうるというセンセーショナルな数字の効果によって、二重構造説は埴原が最初に唱えたように思われていますが、在来の集団に大陸から渡来した集団が混血して日本人が成立するという二重構造説の枠組みは、説明したように明治以来、多くの研究者によって唱えられてきましたし、埴原が唱える直前にも人類遺伝学の尾本恵市や形質人類学の池田次郎、山口敏といった研究者によっても提唱されていました。
二重構造説の問題点
二重構造説が唱えられた1990年代の初頭までは、人類の起源に関しては、「現生人類は各地域の原人が独自に進化して成立した」とするいわゆる「多地域進化説」が人類学の分野の定説でした。しかし、アフリカでの化石人骨の再検討とともに現代人のDNA分析が進んだことで、現在では、私たち現代人はおよそ20万年前にアフリカで誕生し、6万年ほど前にアフリカから出て世界に広がった人々の子孫であるという「新人のアフリカ起源説」が定説として受け入れられています。
この学説の変化は、多地域起源説に従って、原人段階までさかのぼる長いタイムスパンのなかで考えられてきた日本人の起源についても、大きな変更を迫ることになりました。これまでは日本人の起源の問題は、北京原人などに代表される東アジアの原人が、いつ、どのような過程を経て新人となったのか、あるいは東アジアで生まれた新人が、どのような経路で日本列島に到達し、私たち日本人の祖先となったのか、もしくは日本列島のなかでも原人から新人への進化があったのか、という問いに答える必要がありました。しかしアジアでは原人から新人への移行がなかったとすれば、原人段階から新人への進化を考える必然性はありません。たとえ日本で原人段階の化石が見つかったとしても、それを私たち現代の日本に住む集団の起源に関連させて解釈する必要はないからです。人類進化の学説の変化は、結果として最初の「日本人」の出現を従来よりもはるかに新しい時代に設定し、日本人の成立を、6万年前以降に起こったアジアにおける新人の拡散と移動の一部に位置づけることになりました。さらに言えば、日本人の起源の解読には、アフリカから拡散した人類の長い旅路のシナリオの一部を構成するという視点が必要になったのです。しかし二重構造説は、後期旧石器時代の大陸でのヒトの移動を考えていながら、この部分については化石の証拠がほとんどないために、検証された理論になっていません。
二重構造説は、均一な縄文人社会が、水田稲作と金属器の加工技術をもった大陸由来の集団を受け入れたことによって、本土日本を中心とした中央と、南西諸島・北海道という周辺に分化していくというシナリオです。先端技術を受け入れた中央と、その影響が波及しなかった周辺という見方をしているのですが、果たしてこのような視点で、南北3000キロを超え、寒帯から亜熱帯の気候を含む日本列島・南西諸島の集団の成立を正確に説明できるのか、という問題があります。日本列島には、後期旧石器時代にあたる4万年ほど前にホモ・サピエンスが進入したと考えられていますが、大陸との地理的な関係を考えるとルートとして図6-2に示した3つが想定されます。二重構造説では、それぞれの経路を利用して列島に進入した旧石器人が、縄文時代の中期までには日本列島内部で均一化したと仮定しているのですが、そのプロセスに関しての言及はありません。実際には、中期以降の縄文人の形質が比較的均一で、「縄文人」として括ることができることをその根拠としているのですが、そもそも縄文人的な形質を待った集団が、どの範囲まで分布していたのかもよくわかっていませんし、実のところ形態学的に「均一である」とか「似ている」という概念はあやふやで客観的な基準がありません。比較対象との関係で決まりますから、これは弥生時代の人骨との違いが大きいために均一に見えているだけだと捉えることもできるのです。
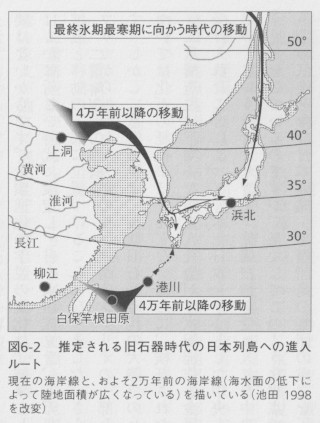
狩猟採集民である旧石器から縄文時代にかけての集団は、その生活が生態環境に依存していたはずです。人々が持つそれぞれの生態系に関する豊富な知識が、そこでの生活を可能にしていたはずですから、異なる
生態系への適応は難しかったと考えられます。旧石器時代の北海道は寒帯から冷温帯に属し、針葉樹林や草原が卓越していました。本州の北半分は冷温帯の針葉樹と落葉樹林、南半分は温帯の針葉樹と落葉樹が広がっており、琉球列島は温暖帯の照葉樹林でした。このような多様な生態環境のなかで生きた人々が均一化に向かうとは考えにくく、むしろ環境の違いは集団の分化を促したと考える方が自然でしょう。列島集団の成り立ちに関して、本州・四国・九州とは生態環境の大きく異なる琉球列島と北海道を個別に考えることが必要です。実際、本土日本、アイヌ、琉球の3集団のミトコンドリアDNAハプログループ頻度は互いに異なっており、特にアイヌと琉球集団の間に類似性は認められません。DNAデータは二重構造説を支持していないのです(図6-3)。
1990年代以降に行われた人骨の形態学的研究では、形態小変異の出現頻度でも琉球列島集団は弥生時代以降の本土日本人に類似し、アイヌとの共通性は認められないこと、アイヌ集団はおおむね縄文人に似ているとしているものの相違点も存在することなどが指摘されています。DNA研究だけではなく形態学の分野でもデータが蓄積されるにつれて、二重構造説では説明のできない事象が報告されるようになっています。そのため、このような単純な捉え方では列島集団の多様性を説明できないと考える研究者も増えつつあります。
縄文人の核ゲノム解析
本書の冒頭で、核のゲノムまで解析するとその個体について何がわかるのかを、北海道の船泊遺跡から出土した縄文人を例に解説しました。ここではゲノム情報を使った集団比較について説明します。分析の手順としては、まず縄文人から抽出したDNAを材料に、次世代シークエンサを使ってそのDNA配列をすべて読み取ります。DNAの状態が良いと、船泊の女性のように現代人と同じ精度でゲノムを決定できますが、さすがにそこまで状態の良いサンプルは滅多になく、たいていはゲノムの数%が解読できるだけです。しかし、そのなかにあるSNPの情報を集めて、他の集団と比較することは可能です。
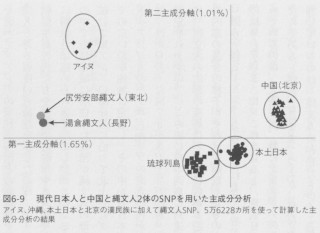
図6-9は現代の日本人と、北京の漢民族と、2体の縄文人のSNPデータを用いて、集団の関係を図式化したものです。膨大なSNP情報を可視化するために「主成分分析」という方法を用いています。この方法は二次元に情報を集約するためにかなりの情報をそぎ落としてしまうのですが、集団間の関係について大まかな傾向を知ることはできるので、この分野の研究によく用いられています。先に紹介した現代日本人のゲノム解析でも使われていました。
縄文人は現代日本人や漢民族集団とは異なる遺伝子構成をしていることがわかるでしょう。これは縄文人の祖先集団が他の集団と非常に古い時代、恐らく東アジアにホモ・サピエンスが進出して間もない時期に分かれた可能性があることを示しています。
全ゲノムを解析した船泊の縄文人女性のデータを使って、アジアの現代人集団との類縁性を調べてみました。その結果、現代人の集団でこの縄文人とある程度の近縁性を示したのは、アイヌ、琉球、本土日本という日本列島の集団に加えて、朝鮮半島の人たちや台湾や沿海州やカムチャッカの先住民でした。一方、いわゆる漢民族との間に類縁性は認められませんでした。このことは、東南アジアから初期拡散によって北上した集団の中で沿岸地域に居住した集団が縄文人の母体になった、と考えると説明がつきそうです。初期拡散で東アジアの海岸線に沿って北上したグループが、台湾付近からカムチャッカ半島に至るまでの広い沿岸地域に定着し、その中から日本列島に進出する集団が現れたのでしょう。ミトコンドリアDNAのハプログループの成立年代からは、縄文人につながる人たちの日本列島への進出は、西から入ったM7aが3万~2万年前、北からはやや遅れてN9bが2万年前以降だったと推定されますので、この頃に日本列島に到達した人々が後の縄文人の母体になったと考えられます。縄文人の内部の変異は、南北に広がる沿岸の各地から日本列島に人々が流入したためと、列島内部の混合の様子がそれほど徹底したものではなかったためであると考えられます。
図6-12(181ページ)にこれまで分析した古代人と現代のアジア集団を併せた主成分分析の結果を示しました。これを見ると、現代日本人が、東アジアの集団の中で図のような位置を占めているのは、大陸集団、特に北東アジアの集団が列島に進入して在来の縄文系集団と混合したためであると解釈できます。つまり、私たち現代日本人の遺伝子の構成をアジアの集団から区別しているのは、縄文人の存在だということになります。一方、私たちのDNAに占める縄文人の割合はそれほど大きくはありません。船泊の縄文人を使って計算すると10%程度になります。ただし、すでに説明したように、私たちにDNAを伝えているのは主に西日本の縄文人である可能性が高いですから、そちらを基準にすればもう少し大きな値になるのでしょう。Y染色体のDNAで縄文人由来と考えられるハプログループDを持つ男性が3割程度、ミトコンドリアDNAの場合は、M7aやN9b、D4b2などを合計すると全体に占める割合は2割程度にはなりそうですから、10%は少し低い値のように思えます。
昨今の縄文土器や土偶を展示した展覧会が多くの人々を集めているのを見ると、日本人のなかには縄文に郷愁を感じる人が多いということを実感します。その根底には、私たちの祖先でありながら失われてしまった人々への想いがあるのだと思いますが、実際に私たちにDNAの大部分を伝えているのは、次に説明する弥生時代になって日本にやってきた人々です。
多様な弥生人
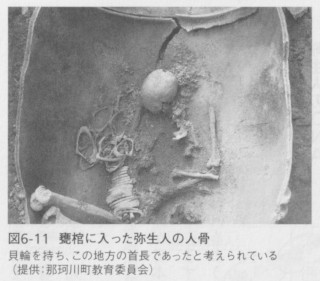
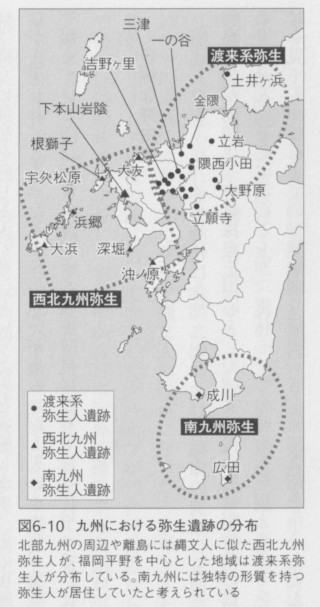 前述したように、弥生時代は北部九州における稲作の開始によって定義されているのですが、列島全体が同時に、狩猟採集を中心とした縄文時代から弥生時代の農耕社会に移行したわけではありません。そしてこのことが、実体としての弥生人を捉えることを難しくしています。九州だけを見ても、これまでの形態学的な研究から、北部九州を中心とした渡来系弥生人、長崎県の沿岸部や離島から出土する、縄文人の直系の子孫と考えられる「西北九州弥生人」、そして鹿児島の種子島の広田遺跡などから出土した、特異な形質を持つ「南九州弥生人」が区別されています(図6-10)。そのようななかで、福岡県や佐賀県などの北部九州地方を中心とした地域では、渡来系弥生人が遺体を甕棺という巨大な素焼きの甕に入れて埋葬する風習を持っていたために、人骨が消失せずに発見される可能性が高く、これまでに数千体の人骨が見つかっています(図6-11)。実際、日本中で発掘された弥生人骨の大部分はこの地域から出土したものなのです。
前述したように、弥生時代は北部九州における稲作の開始によって定義されているのですが、列島全体が同時に、狩猟採集を中心とした縄文時代から弥生時代の農耕社会に移行したわけではありません。そしてこのことが、実体としての弥生人を捉えることを難しくしています。九州だけを見ても、これまでの形態学的な研究から、北部九州を中心とした渡来系弥生人、長崎県の沿岸部や離島から出土する、縄文人の直系の子孫と考えられる「西北九州弥生人」、そして鹿児島の種子島の広田遺跡などから出土した、特異な形質を持つ「南九州弥生人」が区別されています(図6-10)。そのようななかで、福岡県や佐賀県などの北部九州地方を中心とした地域では、渡来系弥生人が遺体を甕棺という巨大な素焼きの甕に入れて埋葬する風習を持っていたために、人骨が消失せずに発見される可能性が高く、これまでに数千体の人骨が見つかっています(図6-11)。実際、日本中で発掘された弥生人骨の大部分はこの地域から出土したものなのです。
すでに説明しましたが、縄文人は上下に寸が詰まって幅が広く、眼窩と呼ばれる眼球を入れる部分が四角い形をしています。そして眉間や鼻骨の隆起が強くて全体的に立体的な顔立ちをしています。平均身長は男性で158センチ、女性で148センチ程度と低いのですが、西北九州の弥生人は、ほぼこのような姿形をしているので縄文人の系統の人々であると考えられています。このような弥生時代にあっても縄文人の形質を残した人骨は、北部九州の沿岸地域、平戸や五島列島の遺跡で発見されています。代表的な西北九州弥生人の遺跡は、佐賀県の玄界灘に面した大友遺跡です。この遺跡では、支石墓と呼ばれる墓に人々が埋葬されているのですが、支石墓は同時期の朝鮮半島南部に多く見られるもので、北部九州と朝鮮半島の関係を示すものと考えられています。となるとこのお墓に埋葬された人たちは渡来系の弥生人だったと考えたくなるのですが、形態学的な研究からは、この遺跡に埋葬された人々は、在来の縄文人に似た人々なのです。
一方、北部九州の弥生人は平均身長で男女とも縄文人よりも5センチほど高くなります。また顔貌ものっぺりとした面長で、鼻根部は平坦です。縄文人とはかなり違った姿形をしているので、両者は由来を異にする集団だと考えられています。朝鮮半島や中国の江南地方から水田稲作をもたらした人たちだと考えられていますので、渡来系弥生人と称されています。
南九州の弥生人は、前述した種子島の広田遺跡から出土したものが有名です。鼻根部とその周辺は縄文人的ですが、顔はさらに上下に短く、後頭部が扁平な特異な形状をしています。身長はさらに低くなり、男性で154センチ、女性では143センチほどだと言われています。九州では、同じ弥生時代にこの3つのタイプの弥生人が同居していたと考えられているのです。
弥生時代が、日本列島の基層集団である縄文人の世界に、大陸由来の集団が流入する時代であったことを考えると、この時期が日本列島においてもっとも遺伝的に多様な集団が分散して居住していた時期だということになります。形質の多様性はそれを反映したものなのでしょう。しかし、弥生人のゲノム解析が行われるようになると、この図式はいささか単純過ぎることもわかってきました。
縄文人と弥生人の混合のシナリオ
先にも説明しましたが、弥生時代は北部九州地域で在来の縄文人との混血が始まり、遺伝的に分化していた集団が均一化に向かう時期でした。そこで誕生した本土の現代日本人の祖先集団は、やがて水田稲作の拡大とともに東進を開始し、在来集団を巻き込みながら東北地方まで進んでいくことになったと考えられます。そのため現代日本人には関西以西の縄文人のDNAがより高い頻度で伝わっている可能性が高いことが予想されるということを指摘しました。このシナリオを元にして、これまでに判明している縄文人のミトコンドリアDNAハプログループの東西の頻度の違いと、本土の現代日本人のハプログループ頻度データを使うことで、東北に到達した段階でどのくらいの人口比があると両者の混合で現代の本土日本人の比率になるか、を推測することができます。
詳しい説明は省きますが、九州歯科大学の飯塚勝さんと奈良女子大学の富崎松代さんにお願いして、モデルに基づくシミュレーションをしてもらいました。西日本縄文人のデータが少ないので、あくまでも現時点で利用できるデータからの類推に過ぎないのですが、人口増加率などのいくつかの仮定をおいて計算すると、西日本系の混合集団が東北の縄文系集団に出会ったとき、数十倍の人口をもって混合が行われたと仮定すると、現代人のハプログループの値になることが示されました。恐らく狩猟採集民である縄文人は、数十人の規模の集落で暮らしていたと考えられるので、そこに流入する稲作農耕民は数百~1000人程度ということになります。列島の内部では、この程度の人口規模の異なる集団が出会い、やがて縄文系の人々が農耕集団に吸収されていったというシナリオが見えてきました。
ただし、このシミュレーションは混合の最初と最後の段階を見ているだけなので、途中の地域がどうであったのか、弥生以降の大陸集団の流人をどう見積もるか、などの検討はできていません。事実、北部九州で始まった稲作は近畿地方には紀元前7世紀に到達しますが、関東には紀元前3世紀にならないと現れません。一方、東北には紀元前4世紀に出現するので、関東よりも早いのです。稲作の東進のルートも単純なものではなかったようなので、信頼性のあるモデルを作るためには、さらにデータが必要です。
私たちの研究グループが最近手がけている、鳥取県の青谷上寺地遺跡から出土した人骨のミトコンドリアDNAハプログループは、遺跡が弥生の後期のものであること、北部九州からは離れた山陰地方にあることなどから、縄文系のハプログループをかなり含んでいると予想していたのですが、解析してみると、ほとんどが弥生時代になって日本列島にもたらされた系統であることが明らかになりました。どうも渡来系弥生人集団の東進もそれほど単純なシナリオに沿ったものではなかったようです。しかし、各地の縄文人と弥生人のDNAデータが揃ってくると、さらに精度の高い予測も可能になるでしょう。このシミュレーションから導かれた集団移動のシナリオを考古学的な証拠とあわせて考えることで、新たな日本人形成のプロセスが見えてくるはずです。
かつては形態の類似度から、弥生時代以降に大陸からの大規模な渡来を予測した研究もありました。その根拠は、縄文人と渡来系弥生人の形質を比較すると、圧倒的に渡来系弥生人の方が現代日本人に似ていることにありました。これに対して考古学の研究者から強い批判が出ました。北部九州の弥生早期の遺跡から出土する朝鮮半島系の土器は、全体の1割程度だと言われており、しかもそれらが出土するのは玄界灘に面した大きな遺跡からだけで、大部分の弥生早期の遺跡には朝鮮系の土器はないのです。これらの事実から、考古学者は弥生時代早期の渡来人の数を、全体の1割程度と見積もっていました。基本的には多数を占める縄文人の血を引く在来系の住民が、水田稲作農耕と金属器という大陸の文化を受け入れたと考えていたのです。
しかしその後、中橋孝博さんと飯塚勝さんによる人口のシミュレーション研究によって、農耕民である弥生人の人口の増加率が、狩猟採集民である縄文人よりも高いことを仮定すれば、最初の渡来者が少数でも数百年で在来系の集団を数の上で凌駕することが示されました。世界中の先住民社会の研究で、一般に狩猟採集民よりも農耕を受け入れた集団の方が、人口の増加率が高いことが示されているので、この仮定には充分な根拠があります。
さらに、弥生時代の開始期が従来の説よりも500年ほどさかのぼることになったことで、渡来系弥生人の人口増加率をさらに低く見積もっても、狩猟採集集団の人口を上回ることになりました。中橋、飯塚の研究によって、多量の渡来人の流人を仮定しなくても、これまで発見された弥生時代前期後半以降の1万基以上の甕棺のなかに残された人骨が、おしなべて渡来系弥生人の形質をしていることを説明できることが示されたのです。しかし、実際の混合の状況はDNAデータを使わないと、精度の高い予想はできません。この分野の研究は、今後どれだけ質の高いDNAデータを集積できるかにかかっています。
北海道先住民の成立史の終わり部分
古代のDNAや骨形態を調べる人類学の研究では、とりあえず人骨がなければ研究が進みません。現在の研究者は各地の博物館や大学に収蔵されている人骨を利用して研究ができますが、私たちの先達は人骨の収集から始める必要がありました。明治、大正、昭和、平成の各時代を通じて、人類学者は各地で人骨の収集に励みましたし、現在でもそれは続いています。その努力なしには人類学の研究は進みません。ただし、過去における人骨の収集方法や、その後の管理には今の基準で考えて適切ではないものもありました。特にアイヌの人たちの人骨の収集に関しては、時代的な背景を考えると無理もない面もあるにせよ、率直に言って私たち人類学者には反省すべき点があります。そしてそのことが原因となって、アイヌの人たちの成立の歴史に関する人類学的な研究を進めることが困難になっている現状があります。ここで紹介した北海道では、縄文時代からミトコンドリアDNAがどのように変遷していったのかを明らかにした研究は、北海道アイヌ
協会の理解を得て行ったものです。人類学の研究は、文献的には13世紀までしかさかのぼることのできないアイヌの人々の起源について、彼らが北海道の縄文人につながる先住民族であることを明らかにしてきました。また、DNA研究はさらに詳細な集団の変遷のシナリオを明らかにしつつあります。これらの研究は、アイヌ民族のアイデンティティに関して重要な貢献ができたと考えています。今後とも、アイヌの皆さんとの協調関係を保ちながら、北海道の集団の成立に関する研究が進むことを願っています。
国家の歴史を超えて
本書ではDNA研究が明らかにした人類の足跡をたどってきました。およそ20万~15万年前にアフリカで誕生した私たち人類すべての祖先が、どの時期にどのようなルートで世界の各地に拡散したかを概観し、最後に、私たち日本人の成り立ちについて見ていきました。その意味では、本書は人類の歴史について書かれた書物ということになります。一般に歴史は、有名な個人や一族、あるいは王権や政権に起きたできごとを中心に語られるものですが、DNAが明らかにする歴史には、基本的には特定の人たちの話は出てきません。DNAの物語る歴史は、個人が持つDNAに刻まれた人類の歩みを手がかりに話が組み立てられていますから、必然的に私たち人類すべてが歩んできた道、日本人すべての成り立ちの物語となるのです。
日本について記述する部分に関しても、この歴史の物語は大部分が日本という国家が成立する前の時代を記述したものです。歴史は国の始まりから説き起こされるのが普通ですから、その点でもずいぶん異色なものになっています。日本に国家が成立したのを1500年くらい前と仮定しても、その頃までには大陸からの大規模な移入も終息し、それ以降に日本の人口比率を変えるほどの大量の渡来があったという事実はなさそうです。ですからDNAに関して見れば、私たちは日本という国ができる前にその材料がそろっていたことになります。この列島に、ある程度の人数が居住し始めた縄文時代以降、国家が成立するまでの期間は、その後の歴史時代の10倍もの長さがあります。私たちのDNAは、その長い時間のなかで、いろいろな地域から流入してきたのでしょう。縄文人が持つDNAは、今では世界中を見わたしても存在しない特殊なものでした。そして弥生時代以降における大陸からの渡来民は、縄文時代に蓄積したDNAのプールに特に大きな影響を与えました。本土日本の集団は、この弥生時代以降に渡来した集団と在来の集団の混血によって成立していったのです。
ある程度地理的に隔離された北海道と沖縄では、本土の日本とは異なる集団の歴史があります。それは、両者が本土日本とは異なるDNAの組成を持っていることからも明らかです。日本列島における集団の成立の歴史は、重層的で複雑なものであることを、私たちの持つDNAは教えています。
国家としての日本は、鎌倉時代の二度にわたる元寇と第二次世界大戦後のアメリカによる占領を除けば、ほとんど他国の侵略や征服を受けることなく、おおよそ単系的に続いてきました。ですから私たちにとって、集団としての日本人がその成立の時点から単系的に続いてきて現在に至るという、いわゆる単一民族であるという考え方は受け入れやすいものです。しかし、ミトコンドリアDNAやY染色体DNAのハプログループを子細に見ていくと、私たちのルーツは大陸の広い地域に散らばっており、それがさまざまな時代にさまざまなルートを経由してこの日本列島に到達し、そのなかで融合していくことによって日本人が成立したことを示しています。実際のところ私たちのルーツを探して時間をさかのぼっていくと、その経路はいくつにも枝分かれし、アジアのさまざまな地域に散らばっていきます。そしてさらに時間をさかのぼっていけば、アジアのなかで複雑に絡み合った道筋が、アフリカに向けて収束していく姿が見えるのです。
もちろん、このような集団の成立史は日本列島のみに固有のものではありません。日本の周辺の地域でも、アフリカから出発した人類につながるさまざまな枝が、同じように混じり合い、それぞれの地域に特有の集団を構成していったのです。特に日本列島へのDNA流入の最大の通路であった、朝鮮半島から大陸の東北地域には、アフリカから出てこの地に至るまでの歴史を、私たちと共有している人々が住んでいます。
私たちはしばしば国の成立と、集団としての日本人の成立を同じものと見なすことがありますが、このように見ていけば、両者は分けて考えるべきものであることがわかります。言うまでもないことですが、日本という国ができる以前に、日本列島には人々が住んでいました。人がいて国ができたということは、国というもののありようを考えるときに、大切な認識だと思います。そして私たちの直接の祖先である人々と、親戚に当たる人たちの子孫が日本の周辺には住んでいます。とかく国同士の関係は、近いところほど複雑になるのですが、同じような道をたどってアフリカからやってきた人々ですから、本質的な違いはないと考えることもできると思います。
家系とDNAのアナロジー
DNAと系統の問題を考えるとき、大きく2つの立揚があります。ひとつは、DNAを変化することなく祖先から連綿と受け継がれる情報として捉える考え方で、もうひとつは自分の持つDNAが多くの人に共有されていると考える立場です。本書の最初にも書きましたが、前者はDNAを血統とか家系と結びつけて捉える考え方で、場合によっては特定の家系を特殊なものであると考える際の生物学的なバックボーンとして利用されることもあります。私たちは家系というものに特別な感情を持っています。これは日本に限らず多くの社会で見られるものですから、人類が共通で持っている考え方の癖のようなものなのかもしれません。ですから生物が祖先から受け継ぎ、子孫へと伝えている遺伝子の正体がDNAであるとわかったとき、家系とDNAを同じようなものとして捉える考え方が生まれました。私たちの持つひとつひとつの遺伝子が、人類進化の過程のどこかの段階で誕生したことは間違いありませんし、それぞれに私たちすべての人類の
共通祖先が存在しますから、これを家系のアナロジーと捉えることも可能です。しかし、私たちは2万個以上の遺伝子を持っているのですから、自身のなかに多数の複雑な系統を持っていることになります。単一の遺伝子の系統をもって、全体の由来を代表させることはできないのです。
先に説明したように、大部分の核DNAは組み換えによって伝わりますから、その系統をたどることは事実上不可能です。しかもDNAは、両親から半分ずつを受け取り、子供に自分のDNAの半分を受け渡すという様式を取るのですから、ひとつの遺伝子に注目すれば、一本道で系統が続くように見えても、遺伝子を総体(ゲノム)で見た場合には、個人の歴史をさかのぼって収束していく道筋のようなものはありえません。むしろ自分の持っている個々の遺伝子が祖先の集団のなかに発散する姿が見えてきます。それは子孫に向かっても同じです。
他のDNAと違って、ミトコンドリアDNAとY染色体DNAは組み換えなしに子孫に伝わるので、ヒトの進化や拡散を研究するのに便利です。ですから、私たちは主にこの2種類のDNAを使って人類の拡散の様子を追跡してきました。この単系統に伝わる性質に注目して、これらのDNAをあたかも家系のシンボルのように捉える考え方が世間にあります。特にY染色体は男系に伝わりますから、最近ではこれを男子の系統のシンボルのように取り扱う不思議な議論が見受けられます。しかし、私たちのDNAは全体としてひとりの人間を作るために働いているのですから、特定のDNAだけをもって、あたかもそれだけが重要であるかのように強調するのはおかしな話なのです。
DNA分析による人類の歴史を解説した啓蒙書のなかにも、ミトコンドリアDNAやY染色体のDNAを、家系のアナロジーとして取り扱っているものがあります。現在の技術では、個人のミトコンドリアDNAやY染色体のハプログループを調べることは、さほど難しくはありませんから、それらを検査してルーツを探す商業的なプログラムも存在します。伝達の経路がハッキリしているこれらのDNAは、その解析がヒト集団の歴史の解明にいかに有効であるかを教えてくれます。しかしそれらは、あくまで集団の歴史を描くのに有効なのでめって、個人の由来を教えるものではないことを認識しておく必要があります。ミトコンドリアDNAやY染色体のDNAは、私たちの持つDNAのごく一部であり、その由来は自分自身の持つすべてのDNAの出自を示しているわけではないのです。
これからの社会と私たちのDNA
狩猟採集民として出発した私たちの祖先は、最初は緩やかな拡散によって、そのテリトリーを広げていきました。農耕や牧畜がはじまった1万年前以降には新たな移住の波が世界に起こり、それが一段落することで現在につながる地域的な違いが生じました。その後、歴史時代を通じてこの地域差は固定化されていきましたが、大航海時代以降の人類の歴史は、細分化した地域集団の境界を曖昧なものにしていきます。ヨーロッパとアフリカからは大量の人々が新大陸に進入し、そこでは遠い昔にアフリカを出て以来、数万年間出会うことのなかった世界中のDNAが集合しました。近代社会になって、交通の発達とともにヒトの移動には拍車がかかり、今や、程度の違いはあるにせよ世界のどこの地域に行っても、人類の持つほとんどのDNAを見いだすことができるようになっています。
この傾向は、社会の状況が大きく変革することでもない限り、促進されることはあっても停滞することはないでしょう。したがって今後も私たちの社会では、人類が長い時間をかけて蓄積してきた、地域に固有のDNAの組成が解消する方向に進むと考えられます。それは日本でも例外ではなく、数百年というタイムスパンで考えれば、今の私たちとは異なるDNAを持つ日本人が多数を占める日がくるというのも荒唐無稽な話ではないと思われます。歴史的に考えれば、現在は縄文時代から弥生・古墳時代への移行期以来二度目となる、外部からのDNAの流入と国内での均一化が進んでいる時期であるとも捉えられます。一度目は数百年以上をかけての流入でしたが、二度目は目に見えて変化がわかるほど急激なものになりました。Y染色体のDNAなどを見ると、日本の社会は大きな混乱もなく渡来した人たちを受け入れて、新たな社会を作ったようにも見えますが、二度目の今回はどのような経過をたどるのでしょうか。移民の問題は、多くの場合、経済と結びついた議論だけが先行しますが、これだけ急速に事態が進むケースでは、伝統や文化を大切にしながら、どのように新たな社会を構築していくかという、私たちの知恵が試される問題になります。
現在、世界的な規模で起こっているヒトの移動は、経済のグローバル化と相まって、国というもののあり方を大きく変えていくでしょう。その潮流のなかで、私たちが国のあり方に関してどのような選択をするのかは、今後の重要な課題となるはずです。「アメリカ・ファースト」に代表される自国中心主義、戦前へ回帰するような最近の風潮は、もしかするとそれに対するひとつの反応なのかもしれません。しかし、今後ますます進むボーダーレスの社会にあって、普遍的な価値を持たないナショナリズムにこだわって未来があるとは思えません。過去と未来を見わたした長期的な視野に立って考えることが大切でしょう。
私たちはかつて、「恒久の平和を念願し、人間相互の関係を支配する崇高な理想を深く自覚するのであって、平和を愛する諸国民の公正と信義に信頼して、われらの安全と生存を保持しようと決意した」と宣言したことがあります。それから70年以上が経って、この考え方が時代に合わないと考える人が増えてきました。しかし、私たちの持つDNAを研究してみると、そもそも人類の持つDNAの違いはごくわずかであること、そしてその成立の経緯から、私たちの持つDNAはほとんどが東アジアの人々に共有されていることがわかりました。少なくとも私たち自身が公正や信義を重んじているのであれば、人類700万年の歴史から見ればほんの少し前に分かれた世界中の人々や、ほぼ同じ遺伝子を持ち、DNAから見れば親戚関係の集団であるアジアの人々にそれを期待することは、それほど間違った話ではないと思います。DNAが明らかにした、人類集団の成り立ちの真の姿は、この日本国憲法前文の精神の正しさを生物学の立場から裏づけているようにも思えるのです。これからの私たちの社会のあり方は、この精神を否定するところからではなく、ここから出発することが求められているでしょう。どのみち、信頼関係が構築できなければ、人類に未来はないのですから。
![[商品価格に関しましては、リンクが作成された時点と現時点で情報が変更されている場合がございます。]](https://hbb.afl.rakuten.co.jp/hgb/1583936b.847a7192.1583936c.2d77a722/?me_id=1213310&item_id=19555465&pc=https%3A%2F%2Fthumbnail.image.rakuten.co.jp%2F%400_mall%2Fbook%2Fcabinet%2F2553%2F9784140912553.jpg%3F_ex%3D240x240&s=240x240&t=picttext "[商品価格に関しましては、リンクが作成された時点と現時点で情報が変更されている場合がございます。]")
